
 教職員募集
教職員募集 所内専用
所内専用大洋の東西で異なるマイワシの環境応答 ――耳石が示すグローバル生存戦略の鍵――
2022年10月16日
東京大学 大気海洋研究所
水産研究・教育機構 水産資源研究所
京都大学

研究成果
発表のポイント
◆世界の温帯海域に分布するマイワシのなかで、北太平洋の東側(カリフォルニア海流域)と西側(黒潮―親潮域)に生息するマイワシでは、生後数カ月間の生息水温と、成長速度・代謝速度およびそれらの水温変動に対する応答が大きく異なることを発見しました。
◆地球の各海洋の西側のマイワシの個体群が寒冷な年代に、東側に生息するマイワシの個体群が温暖な年代にそれぞれ増大するという長年の謎を説明できる可能性があります。
◆耳石という頭部にある硬組織に最新の化学分析手法を用いることで、魚が経験してきた水温環境やエネルギー消費量などの変化を十日程度の単位で調べることが可能になりました。観測が困難な海の中での仔稚魚の生態を理解することで、様々な水産資源の変動要因が解明されることが期待されます。
発表者
坂本 達也(研究当時:水産研究・教育機構水産資源研究所 JSPS特別研究員 / 現:東京大学大気海洋研究所 海洋科学特定共同研究員 / ポルトガル海洋大気研究所 JSPS海外特別研究員)
髙橋 素光(水産研究・教育機構水産資源研究所 主幹研究員)
小松 幸生(東京大学大学院新領域創成科学研究科 准教授)
白井 厚太朗(東京大学大気海洋研究所 准教授)
石村 豊穂(研究当時:茨城工業高等専門学校 准教授 / 現:京都大学大学院人間・環境学研究科 准教授)
樋口 富彦(東京大学大気海洋研究所 特任研究員)
発表概要
世界中の温帯海域に生息するマイワシの個体数は、環境変動に応答して激しく増減することが知られています。その中で、日本近海のように各大洋の西側に生息するマイワシ個体群は比較的寒冷な年代に増加するのに対し、東側に生息する個体群は温暖な年代に増加することは大きな謎となっていました。水産研究・教育機構、東大大気海洋研究所、国立台湾大学、アメリカ海洋大気庁、京都大学らからなる共同研究チームは、日本近海と北米西岸で収集されたマイワシの耳石(図1、注1)について、日単位での成長解析に加えて、非常に細かい解像度での安定同位体分析(注2)を行いました。その結果、北太平洋の東西の集団間では、初期成長速度、エネルギー消費量、分布水温が大きく異なることが見出されました(図2)。このような東西の対称的な違いは、遠く離れた南アフリカの西岸(大西洋の東側)と南・東岸(インド洋の西側)にいるマイワシ集団の間でも、共通して観察されました。さらに、日本近海のマイワシは稚魚期の終わりまでの成長速度が低水温下で高くなるのに対し、北米西岸のマイワシでは高水温下で高くなることが示されました(図3)。成長が速い仔稚魚は生残率が高くなるため、このように成長速度の水温応答が逆転していることが、世界の大洋の西側と東側のマイワシ個体数の環境応答の違いを生んでいるのではないかと考えられました。こうして多様な環境に適応して生き様を変え、海域により異なる環境応答を示すことが、マイワシの気候変動に対するリスクヘッジになっている可能性があります。従来観測が困難だった海中の仔稚魚の生態を理解することで、様々な水産資源の変動要因が解明されることが期待されます。
発表内容
<研究の背景>
地球上の中緯度帯の海は一般に、暖流が流れる西側で暖かく、寒流が流れかつ深層水が表層に湧き上がる東側で冷たくなります。マイワシ属(Sardinops属)は西大西洋を除く全ての海洋の西・東側両方に分布し、水産資源として漁業を支えるとともに、海鳥、海棲哺乳類、大型魚類など様々な捕食者の餌となる重要な生物です。しかし各海域のマイワシ個体数は数十年程度のサイクルで、1000倍にも及ぶ幅で増減することが知られています。このような大規模な個体数変動は、過去数千年にわたり繰り返し発生しており、様々な気候変動指数と連動していることから、主に環境変動に起因する自然現象だと考えられてきました。しかし、どのような仕組みで気候変動とマイワシの個体数変動が結びつくのか、そのメカニズムについては多くの点で不明です。特に、日本近海を含む各大洋の西側に生息する個体群は比較的寒冷な年代に増加するのに対し、東側に生息する個体群が温暖な年代に増加することは、大きな謎となっていました。
多くの魚類は大量の卵を産みますが、そのほとんどは仔稚魚期を通じて死亡します。過去の研究では、魚類では一般に集団中で極めて成長速度が高かったものだけが生き残ることや、マイワシでは稚魚期の成長が速い年には生き残った個体が多かったことが報告されてきました。従って、環境変動が成長速度に与える影響を知ることが、仔稚魚期の死亡率や集団の個体数変動の仕組みを理解することにつながります。一方で、広大な海の中で、マイワシの仔稚魚がどこを泳ぎ、どのような環境を経験したかを知ることは困難であり、環境変動と成長の関係の理解は進んでいませんでした。
<研究内容>
本研究では、魚類の内耳にある炭酸カルシウムの結晶である耳石(図1)について最新の分析を行うことで、この困難を乗り越えました。マイワシの耳石は毎日1本の成長輪を形成しながら成長し、耳石の大きさは体長と強く相関することから、日々の成長によるサイズ変化の履歴を推定することができます。さらに、耳石の酸素・炭素安定同位体比(注3)からはそれぞれ、周囲の水温と魚のエネルギー消費量を推定することが可能です。これまでの同位体分析は解像度が低いことが難点でしたが、研究グループが進めてきた技術開発により、10日程度の解像度で分析可能になりました。この技術を、2006-2010、2014、2015年の秋に北海道東部沖合域から採集された日本のマイワシ(Sardinops sagax melanostictus)および1991-1998、2005-2007年に北米西岸の南カリフォルニア湾沿岸域で採集されたカリフォルニアマイワシ(Sardinops sagax sagax)の耳石に用いることで、野外環境における仔稚魚期の生息水温、成長率、エネルギー消費量のセットを世界で初めて解析しました。
まず、孵化から4-5カ月の間、成長速度、代謝速度、経験水温は全て、日本のマイワシが北米西岸のマイワシを上回ることがわかりました(図2)。成長速度については、遠く離れた南アフリカ海域のマイワシとも比較したところ、日本のマイワシの成長履歴は南アフリカ南・東海岸(インド洋の西端)のマイワシのものと類似し、北米のマイワシの成長履歴は南アフリカの西海岸のマイワシ(大西洋の東端)のものと類似していました。これらのことから、海洋の西側と東側とで大きく異なる海洋構造・海洋環境が、マイワシが仔稚魚期に経験する水温、エネルギー消費量、成長速度に強く反映されていることが示されました。
さらに、水温変動と成長速度の関係は日本と北米西岸のマイワシで異なることがわかりました(図3)。日本のマイワシでは、仔魚期は暖かい水温下で成長速度が高くなりましたが、年齢とともにその関係は変化し、生後3カ月半の稚魚は経験水温が低いほど体長が大きい傾向にありました。一方、北米のマイワシは、仔稚魚期を通じて、経験した水温が高いほど体長が大きい傾向がありました。水温とエネルギー消費量の関係から求めた活動に最適な水温(注4)との比較から、日本のマイワシでは水温による生理状態への影響が、北米のマイワシでは水温変化に伴う周囲の餌の量や酸素濃度などの環境変化の影響が、成長速度を支配的に規定していると考えられました。高い成長速度は捕食による死亡率を低下させるため、このように太平洋の東西で水温変化と成長速度の関係が逆になることが、それぞれのマイワシの個体数が寒冷期/温暖期に増大する理由になっていると考えられました。東西太平洋と南アフリカのマイワシの生息環境や生活史特性が類似していたことから、同様のメカニズムが世界の海の西側と東側のマイワシで普遍的に機能している可能性があります。
地球温暖化をはじめとする将来の気候変動に対して、海の魚の数がどう変化していくのかを予測することは、依然として困難です。しかしマイワシ属は、海の西側・東側のように大きく異なる環境に適応し、環境変化に対してそれぞれ異なる方向に成長速度などの生活史特性が変化するため、将来の気候変動がマイワシ属全体の存続に与える影響は小さいと考えられます。多様な環境に適応し、環境変化に対しても多様な応答を示すことが、予測困難な気候変動に対するリスクヘッジになっており、それこそがマイワシの進化戦略の鍵なのかもしれません。このように、新しい技術を開発し、野外生物の生態の理解を一歩一歩進めていくことが、人間社会が生態系との間に持続可能な関係を築くために重要であると考えられます。
本研究は、科研費(17J00556、22780185、22H05026、 22H05028、16H02944、18H04921)の支援により実施されました。また本研究は、水産庁水産資源調査・評価推進委託事業による成果の一部を活用しています。
発表雑誌
雑誌名:「Nature Communications」(10月16日付)
論文タイトル:Contrasting life-history responses to climate variability in eastern and western North Pacific sardine populations
著者:Tatsuya Sakamoto*, Motomitsu Takahashi, Ming-Tsung Chung, Ryan R. Rykaczewski, Kosei Komatsu, Kotaro Shirai, Toyoho Ishimura, Tomihiko Higuchi
DOI番号:10.1038/s41467-022-33019-z
アブストラクトURL:https://doi.org/10.1038/s41467-022-33019-z![]()
問い合わせ先
東京大学大気海洋研究所 海洋化学部門 海洋科学特定共同研究員 / ポルトガル海洋大気研究所 JSPS海外特別研究員
坂本 達也(さかもと たつや)
E-mail:tatsfish◎g.ecc.u-tokyo.ac.jp ※「◎」は「@」に変換してください
東京大学大学院新領域創成科学研究科 准教授
小松 幸生(こまつ こうせい)
E-mail:kosei◎aori.u-tokyo.ac.jp
用語解説
- 注1:耳石
- 魚類の内耳の中で形成される、炭酸カルシウムを主成分とする結晶です。毎日輪紋を刻みながら少しずつ大きくなり、その化学成分は周囲の環境や魚の代謝量によって変化します。輪紋の数を数えることで日齢・年齢を知ることができるほか、化学分析によって魚の経験した環境を調べることができるため、「魚の日記」と言われることもあります。
- 注2:安定同位体
- 同じ元素でありながら、わずかに重さの異なる原子のことを同位体といいます。その中で、放射性崩壊によって他の原子に変化することなく安定して存在するものを安定同位体比といいます。例えば今回注目した酸素原子では、原子の重さの指標となる質量数が16のものが自然界の大半を占めますが、17や18のものもわずかに存在します。
- 注3:耳石の酸素・炭素安定同位体比
- 耳石の酸素安定同位体比は、魚の周囲の水温が高いと低くなる(耳石の中の質量数18の酸素原子の割合が減る)性質があるため、経験した水温の指標となります。また、耳石の炭素安定同位体比(質量数13の炭素原子の割合)はエネルギー消費量の指標になります。これは耳石中の炭素には、環境から取り入れた二酸化炭素と魚自身が呼吸をすることで発生した二酸化炭素の2種類のソースがあり、魚の呼吸量が増えると後者の割合が増加するためです。一般に、呼吸由来の炭素の同位体比は環境由来の炭素の同位体比と比べて著しく低いため、耳石の炭素安定同位体比も魚のエネルギー消費が激しいと低下することになります。
- 注4:活動に最適な水温
- 生物のエネルギー消費量は、下限値と上限値がそれぞれ独立に水温によって規定されています。下限値は、魚が全く動かず餌も一切食べないときに、体の機能を維持するためだけに消費するエネルギー量、上限値は魚が限界まで活動する際のエネルギー消費量です。生物はこの下限値と上限値の差が最大になる温度において活動に最も多くのエネルギーを使うことができるため、それが活動に最適な水温となります。野生個体の実際のエネルギー消費量も、下限値と上限値の幅が最大化する最適な水温の近くで、最も多様な値を示すことが期待されます。ここから、野生個体のエネルギー消費量の値の幅が最大になる温度を、活動に最適な水温として見積もりました。
添付資料

図1.マイワシの耳石(左)、耳石の中心部拡大図(中央)、耳石を削る精密マイクロドリル(右)
耳石は1日1本輪紋を刻みながら成長する(中央)。耳石をマイクロドリルで削り(右)、得られる微量の粉末の安定同位体比を分析した。京都大学・石村准教授が開発した超微量分析システム(MICAL 3c)は、世界で最も少ない量で高精度の分析ができる。
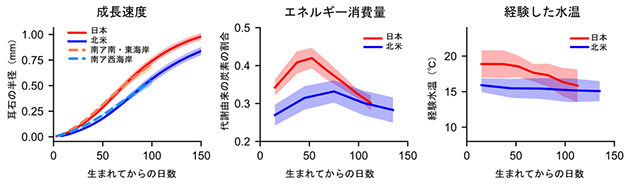
図2.北太平洋、南アフリカ沿岸のマイワシの成長速度、エネルギー消費量、経験水温の違い
成長速度は、海の西側である日本近海と南アフリカの南・東海岸(インド洋の西端)で速く、東側である北米西岸と南アフリカの西海岸(大西洋の東端)で遅くなっていた。西側で暖かく、東側で冷たいという明確な海洋環境の違いが反映されていると考えられる。
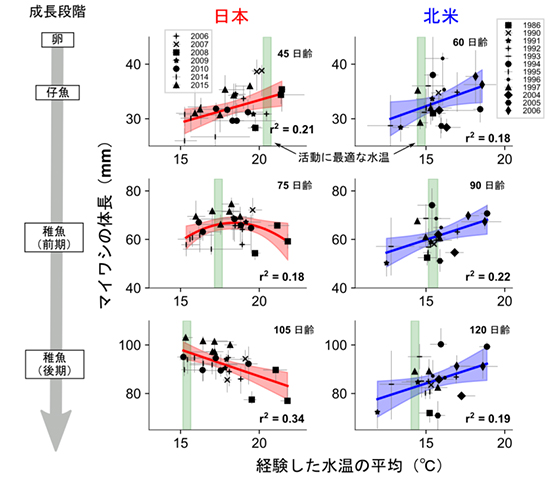
図3.日本と北米のマイワシの水温変化とサイズの関係
日本のマイワシは低水温下で、北米のマイワシは高水温下で、稚魚期の終わりまでの成長が良くなるという違いがあった。
![]()