
 教職員募集
教職員募集 所内専用
所内専用マサバの成長と個体数変動のモデル化を実現
2023年4月12日
東京大学 大気海洋研究所
発表のポイント
◆太平洋のマサバに関して、卵から成魚における成長と個体数変動を表現できる生物エネルギー・個体群動態結合モデルを初めて開発しました。
◆1998年から2018年にかけて21年間の水温および餌料環境を与えた過去再現実験で2010年以降の個体数の増大を再現しました。
◆モデル解析結果から、水温と餌料環境の両者が好適であることに加え、漁獲圧が適切に管理されていることがマサバ個体数増大に重要であったことが示唆されました。
発表者
王 子欽(東京大学大学院農学生命科学研究科 修士課程)
伊藤 進一(東京大学大気海洋研究所 教授)
矢部 いつか(東京大学大気海洋研究所 特任研究員)
郭 晨穎(中国科学院南海海洋研究所 特任研究員)
発表概要
マサバは、太平洋沿岸域に広く分布する重要な水産資源の一つですが、資源量(注1)は気候変動や漁獲圧の影響を受けて変動することが知られています。気候変動は耐性の弱い仔稚魚の生残に強く影響し、漁獲圧は産卵前の成魚の数を減少させることで総産卵量に影響します。東京大学大気海洋研究所の伊藤進一教授を中心とする研究チームでは、マサバの成長-回遊モデルを開発し、過去のマサバの仔稚魚の成長を再現していましたが、その後の成魚までの成長や産卵に関するモデルは存在せず、マサバの個体数(人間の人口に相当)変動を成長、漁獲圧、産卵と同時に解析できるツールはありませんでした。
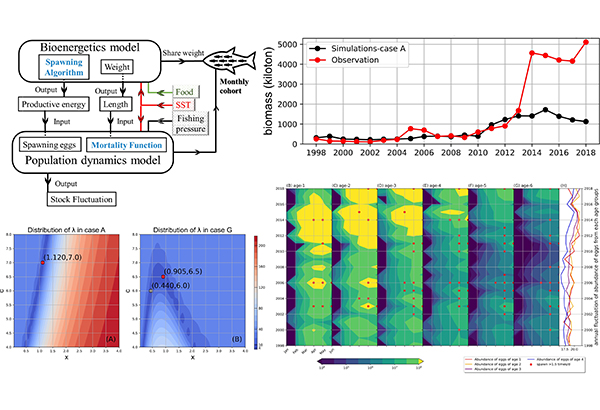
そこで、同研究チームは、マサバの成長を再現する生物エネルギーモデル(注2)を成魚まで拡張し、さらに産卵された卵が仔魚、稚魚へと成長する間の死亡率を体長や成長に依存させて表現する個体群動態モデルと組み合わせることで、生物エネルギー・個体群動態結合モデル(注2)を開発しました。また、成魚の漁獲による個体数減少もモデルで表現できるようにしました。
1998年から2018年にかけて21年間の衛星データから求めた水温および餌料環境を与えた過去再現実験を実施し、2010年以降の個体数の増大には、水温と餌料環境が好適であることに加え、漁獲圧が適切に管理されていることが重要であったことが示しました。
本研究成果は、2023年4月11日(イギリス夏時間)に英国科学誌「Frontiers in Marine Science」のオンライン版に掲載されました。
発表内容
〈研究の背景と問題点〉
マサバは、マイワシ、カタクチイワシ、サンマなどと同様に北西太平洋に生息している多獲性の重要な水産対象種です。しかし、日本の太平洋側のマサバの個体数は、1970年代の増大期、1990年代の減少期、2010年代の増大期と数十年周期で10倍以上の変動を繰り返すことが知られています。先行研究によって、マサバの仔稚魚期の生残が、マサバの個体数変動に重要であることが示されており、仔稚魚期の成長が良いときに生残率が高くなることが知られていました。そのため、マサバの仔稚魚期の成長を再現する生物エネルギーモデルが開発されていましたが、成魚に至るまでの成長を再現するモデルは存在せず、成魚が産卵する卵の数、仔魚、稚魚へと成長する過程の死亡などを加味して、マサバの個体数変動を表現できるモデルがありませんでした。このため、海洋環境変動がマサバの成長を通して、マサバの個体数変動に与える影響を統合的に調べることができず、研究の進展を妨げていました。
〈研究内容〉
本研究では、これまで開発していた太平洋のマサバ仔稚魚の生物エネルギーモデルを成魚にまで拡張し、卵巣へのエネルギー蓄積をモデル化し、産卵量をモデル内で推定できるようにしました。産卵された卵は、水温が高いと早く孵化する効果をモデルに取り入れて、水温変動が卵期の長さに影響することで卵の累積死亡数が変化する効果を表現しました。孵化した仔魚は摂餌によってエネルギーを得ますが、摂餌量は海水中の餌料密度が高いほど高く、また適水温帯に近いほど高くなるようモデル内で表現しました。
先行研究によって、大西洋のタイセイヨウサバの仔魚期における死亡率は体長と成長率に依存することが示されていたため、太平洋のマサバでも仔稚魚期に体長と成長率に依存するとモデル内で仮定しましたが、体長依存性と成長率依存性に関する知見が存在しませんでした。そのため、1998年から2018年にかけて21年間の衛星データから求めた水温および餌料環境を与えた過去再現実験を実施し、計算結果として得られる1998年から2014年のマサバの資源量と再生産成功率(注3)が観測値とあうように最適な体長依存性と成長率依存性を求めました。この際、1998年から2018年までの実際の漁獲圧もモデルで与えました。このようにしてマサバの成長、漁獲、産卵と個体数変動を表現する生物エネルギー・個体群動態結合モデル(図1)を開発し、1998年から2018年までの個体数、資源量、再生産成功率の再現計算を実施しました(図2)。
その結果、マサバの死亡率には、体長依存性が重要であり、成長とともに体長が大きくなることで急激に死亡率が減少するため、成長が早いと仔稚魚期の死亡が抑えられることが示されました。仔稚魚期の成長は、海水中の餌料密度に大きく依存し(図3)、2000年代に入ってから餌料環境が好転したため、マサバ仔稚魚の成長が良くなり、2010年以降の個体数の増加に結びついたことが示唆されました。ただし、卵期の累積死亡数は水温にも大きく依存するため、水温および餌料密度の両者が好適であることが2010年以降の個体数増加にとって重要であったことが示唆されました。
今回開発した生物エネルギー・個体群動態結合モデルには、漁獲によるマサバ個体数の減少も考慮されていますが、漁獲圧を高めると2010年以降の個体数増加が生じなくなることが確認され(図4)、2010年以降の個体数増加には適切な資源管理も重要であったことが示唆されました。
〈社会的意義と今後の展望〉
本研究により、太平洋のマサバの成長、個体数、資源量の変動を統合的にモデリングすることができました。従来の資源管理では、海洋環境の影響を直接取り入れていませんでしたが、今回開発したモデルを用いることで、海洋環境変動がマサバの成長を通して、個体数、資源量に影響する仮定を表現することができます。将来、地球温暖化によって環境変動が激化するなか、海洋環境の影響を加味した資源管理が求められることが予想され、本モデルがその基礎になると期待されます。
現在のモデルでは、マサバの回遊経路が単純化されていますが、今後はマサバの回遊行動を取り入れ、分布域の移動も表現できるモデルへと展開する予定です。
本研究は、文部科学省・科学研究費補助金 基盤研究(A)「サンマ初期生活史の回遊経路の非連続性と分布沖合化維持機構の解明(課題番号:21H04735)」および学術変革領域研究(B)「生物地球化学タグによる回遊履歴復元学の創成(課題番号:22H05030)」の支援により実施されました。
発表雑誌
雑誌名:「Frontiers in Marine Science」(4月11日付)
論文タイトル:Development of a bioenergetics and population dynamics coupled model: A case study of chub mackerel
著者:Ziqin Wang, Shin-ichi Ito*, Yabe Itsuka and Chenying Guo
DOI番号:10.3389/fmars.2023.1142899
アブストラクトURL:https://doi.org/10.3389/fmars.2023.1142899![]()
問い合わせ先
東京大学大気海洋研究所 海洋生命システム研究系 海洋生物資源部門
教授 伊藤 進一(いとう しんいち)
E-mail:goito◎aori.u-tokyo.ac.jp ※「◎」は「@」に変更してください
用語解説
- 注1:資源量
- ある魚種の総個体の体重を積算した全重量を資源量と呼ぶ。
- 注2:生物エネルギー・個体群動態結合モデル
- 魚類の成長を、摂餌によって獲得したエネルギーと、呼吸代謝、消化、排出、排泄などの消費エネルギーの差によって計算するモデルを生物エネルギーモデルと呼ぶ。また、魚類の個体数の変動を捕食者からの被食による自然死亡率や漁獲による死亡、産卵による増大などによって計算するモデルを個体群動態モデルと呼ぶ。両者を結合し、成長と個体数変動を計算できるようにしたモデルを、生物エネルギー・個体群動態結合モデルと呼ぶ。
- 注3:再生産成功率
- 親魚資源量あたりの加入個体数を再生産成功率と呼ぶ。1998年の再生産成功率は、1998年に生まれたマサバのうち翌年に生き残ったマサバの個体数を、1998年に産卵可能だった親魚の資源量で割り算することで求める。
9.添付資料
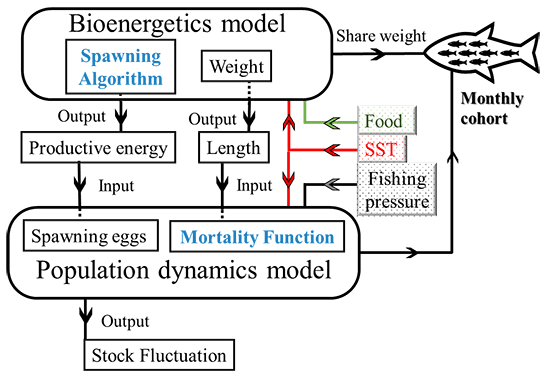
図2 1998年から2018年の(A)個体数、(B)資源量、(C)再生産成功率の観測値(黒線)とモデル計算結果(赤線)。マサバの個体数などは漁獲量などを参考に推定するが、漁獲対象が7歳魚程度まであるため、すべての年齢時の漁獲統計が整理されていない2015年以降の値は信頼性が下がるため、1998年から2014年の観測値を再現するようにモデルの中の死亡率が調整されている。
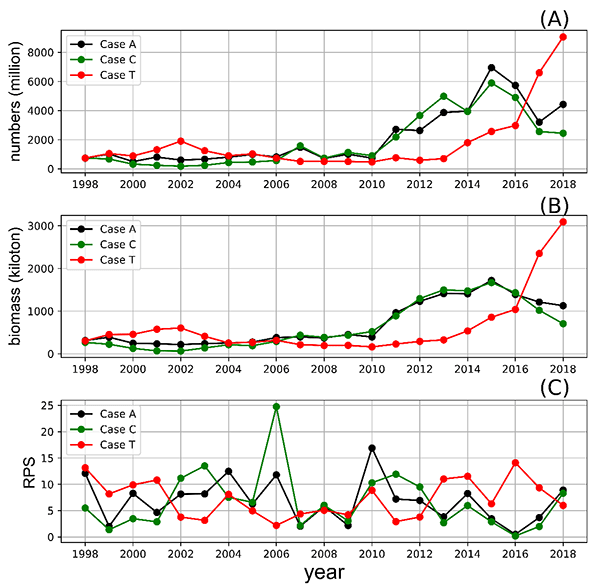
図3 1998年から2018年の(A)個体数、(B)資源量、(C)再生産成功率のモデル計算結果(黒線)と、餌料密度の年々変動のみを与えた計算結果(緑線)および水温の年々変動のみを与えた計算結果(赤線)。餌料密度の年々変動が2010年以降の個体数増大に重要であることがわかる。一方で、2014年以降の個体数増大に水温が大きく影響していることがわかる。
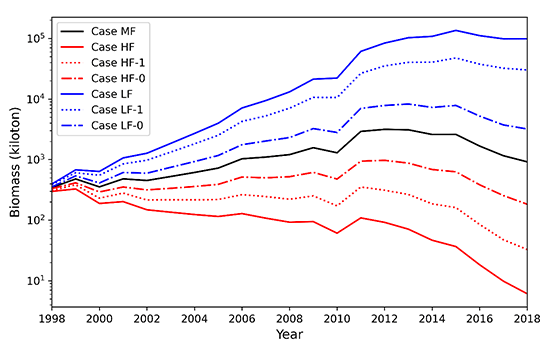
図4 漁獲圧を変化させた場合の1998年から2018年の資源量のモデル計算結果。
MF:全年齢に1998年から2018年の平均漁獲圧を与えた場合
HF:全年齢に平均漁獲圧に1998年から2018年の漁獲圧の標準偏差を加えた漁獲圧を与えた場合
HF-1:1歳魚以上に平均+標準偏差の漁獲圧を与えた場合
HF-0:0歳魚に平均+標準偏差の漁獲圧を与えた場合
LF:全年齢に平均-標準偏差の漁獲圧与えた場合
LF-1:1歳魚以上に平均-標準偏差の漁獲圧を与えた場合
LF-0:0歳魚に平均-標準偏差の漁獲圧を与えた場合
漁獲圧を増やした赤線のケースでは、2010年以降の資源量の増大が抑制されるのがわかる。
![]()
![]()