
 教職員募集
教職員募集 所内専用
所内専用来たのは誰だ 新たに発見されたカワアイ集団の遺伝的特性
2008年4月
生態系動態部門 底生生物分野

最近、様々な生物種で分布が北へ拡大した事例が報告され、地球温暖化の影響ではないかと疑われています。拡大する速度は種によって違うので、生態系を構成する種の組成が変化し、長い時間をかけて培われた種間相互作用のネットワークが破壊されて、生態系の構造に不可逆的な変化がもたらされることが心配されます。私達がこれまで研究してきた干潟の巻貝(小島 2002, 2008)でも今世紀に入ってから、これまで分布が知られていなかった場所で新しい個体群が発見されています(Kojima et al. 2005, 2008; 小島 2007)。また、船舶のバラスト水や海外から輸入されたアサリなどの種苗に混じって日本に侵入する移入種の問題も続いており、中にはサキグロタマツメタの様に、地元の水産業に壊滅的な打撃を与えているものもあります。
カワアイ(図1)は日本の干潟の代表的な巻貝類のひとつでしたが、高度成長期の埋立てや沿岸環境の悪化により、九州より北の地域で絶滅が危惧されています。日本以外に朝鮮半島や中国にも分布しています。私達はミトコンドリアDNAのチトクロームcオキシダーゼ・サブユニットI(COI)遺伝子の塩基配列に基づく解析をおこない、日本列島や韓国のカワアイ集団には、琉球列島の集団の持つ遺伝的多様性のごく一部しかないことを明らかにしました(Kojima et al. 2004)。これは、最終氷期が終わった後に、短期間の内に九州以北に分布域が広がったためと考えています 。

図1a:宮城県万石浦での調査風景

図1b:採集されたカワアイ
東北地方では、最近までカワアイの分布が知られていませんでしたが、2002年と2004年に宮城県松島湾で、2004年に福島県松川浦で、2005年に宮城県万石浦で、東北大学の鈴木孝男博士らによって、新しい集団が発見されました。松島湾では、2002年にイボウミニナの新集団も発見されており、温暖化の影響や人為的な移入の可能性が考えられています。また、90年代に絶滅したと考えられていた東京湾でも2001年に、湾奥部の人工潟湖(新浜湖)でカワアイの繁殖が確認されています(飯島ら 2002)。
これら新発見のカワアイ集団がどこから来たのかを調べるため、松川浦、万石浦、新浜湖でカワアイを採集し、COI遺伝子の塩基配列を日本や韓国の他の集団と比べました。その結果、東北の2つの集団は遺伝的多様性が低いものの、本州西部や九州の集団と統計的に有意な遺伝的差異が認められませんでした。このことから、これらの集団は西日本からの移入又は温暖化等による分布域拡大によるものである可能性が高いと考えられます。一方、東京湾の集団では21個体中9個体が琉球列島も含めて、他の日本や韓国の集団に見られない変異を示しました(Kojima et al. 2008; 図2の黄色)。
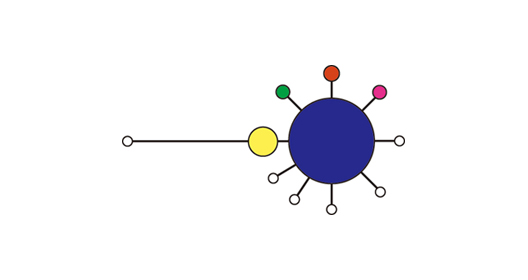
図2a:ミトコンドリアDNAのチトクロームcオキシダーゼ・サブユニットI(COI)遺伝子の塩基配列に基づくカワアイの遺伝子ネットワーク
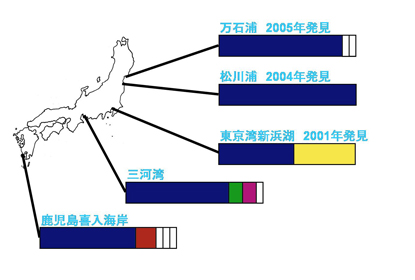
図2b:各地域集団の遺伝的組成 (b). 棒グラフの色は遺伝子ネットワークの各ハプロタイプに対応する
さらに解析を進めるために、遺伝的に異なる本州と沖縄のカワアイ、各1個体についてミトコンドリアDNAの約半分(8500塩基対)の塩基配列を決定し、比較しました。カワアイの遺伝子配列は、これまで軟体動物から報告されているどの配列とも異なっていましたが、近縁なウミニナやホソウミニナについて決定した約4000塩基対に含まれる遺伝子の配列と同じでした(図3)。個体間の変異の大きかった二つの遺伝子領域(NADHデヒドロゲナーゼ2(ND2)遺伝子、12SリボゾームRNA遺伝子と16SリボゾームRNA遺伝子の間の領域)について、COIの場合と同様の解析をおこいました。その結果、9個体がND2遺伝子に他の集団にない変異を共有しており、そのうち8個体はさらに第3の領域にも変異を共有していました。COI遺伝子と合わせると、21個体中18個体が日本の他の集団にない変異を持っていました。
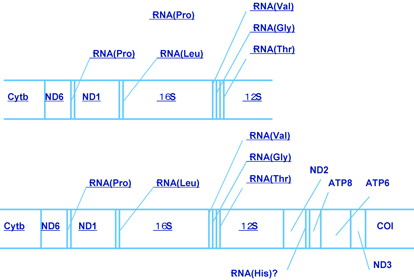
図3: ホソウミニナ・ウミニナ(上)とカワアイ(下)のミトコンドリアDNAの遺伝子配置
以上の結果から、現在の東京湾のカワアイ集団は、本来の東京湾の集団が絶滅した後に、海外からの人為的移入によって新たに形成された可能性が高いと考えられます。新浜湖は東京湾の湾奥に1970年に造成された人造湖です。カワアイが見つかった干潟は1977年ごろできたと言われています。周辺の干潟でアサリ種苗の放流がおこなわれた記録はありませんが、近くの海浜公園では、潮干狩り用にアサリの放流が行なわれています。また東京港に入る外国船のうち、最も数の多いのは中国籍船です。こうした経路で、外国産のカワアイの侵入が起きたのかもしれません。
日本の沿岸には、カワアイ以外にもアジア大陸との共通種が数多く分布しています。その様な種では、海外からの移入があっても、形態での識別することが困難です。知らず知らずのうちに日本の生物集団の遺伝的特性が蝕まれているかもしれません。また今後、温暖化の進行に伴って沿岸生物の分布域拡大や移入種の定着が加速する事も予想されます。定着した個体が、その場所に元々いた集団の遺伝的性質をどの様に変化させるのかを明らかにするため、日本の沿岸生物の集団構造に関する情報を網羅的に収集し、移入が疑われる種について数年程度の時間を置いて調査を繰り返しおこなう必要があると思われます。
参考文献
飯島明子, 黒住耐二, 風呂田利夫. 2002. 東京湾人工潟湖干潟に形成された絶滅危惧種の干潟産腹足類カワアイCerithidea djadjariensis (Martin)(軟体動物門、腹足綱)の個体群. 日本ベントス学会誌. 57: 34-37.
小島茂明. 2002. 干潟の巻貝に記録された海洋環境変動. 遺伝. 56(2): 62‐66.
Kojima S, S Kamimura, A Iijima, T Kimura, K Mori, I Hayashi, and T Furota. 2005. Phylogeography of the endangered tideland snailBatillaria zonalis in the Japanese and Ryukyu Islands. Ecological Research. 20: 686-694.
Kojima S, S Kamimura, A Iijima, T Kimura, T Kurozumi, and T Furota. 2006. Molecular phylogeny and population structure of tideland snails in the genus Cerithidea around Japan. Marine Biology. 149: 525-535.
小島茂明. 2007. 地球温暖化で変わる干潟の巻貝? 干潟ウォッチングフィールドガイド 市川市・東邦大学理学部東京湾生態系研究センター編 誠分堂新光社. 109.
Kojima S, S Ozeki, A Iijima, K Okoshi, T Suzuki, I Hayashi, and T Furota. 2008. Genetic characteristics of three recently discovered populations of the tideland snail Cerithidea djadjariensis (Martin) (Mollusca, Gastropoda) from the Pacific coast of the eastern Japan. Plankton and Benthos Research. 3: 96-100.
小島茂明. 2008. 日本沿岸における底生動物の分散と遺伝的分化. 海洋生命系のダイナミクスシリーズ第5巻「海と生命」,塚本勝巳(編). 東海大学出版会. 印刷中.
![]()