
 教職員募集
教職員募集 所内専用
所内専用マアナゴの産卵場はどこにある?
2008年4月
海洋生命科学部門 行動生態計測分野

寿司や天ぷらで人気の高いマアナゴConger myriasterが,一体どこで産卵しているのか,ニホンウナギと同様に,長い間 謎となっている.これまで産卵場は東シナ海の陸棚縁辺部にあると思われていた (Takai 1959). しかしそこでは,生まれてかなり時間が経った大きな仔魚(レプトセファルス)しか採れていない(Kurogi et al. 2002).マアナゴは水産重要種であるため,その生活史や集団構造を知ることが重要で,これまでにもいくつか研究がある (岡村ら2000; Kimura et al. 2004).しかし生活史研究の中で最重要課題ともいえる産卵場の問題は,なかなか進展が見られなかった.大型のレプトセファルスやその後変態した稚アナゴは日本の沿岸域で容易に採集できるが(図1のd-f),これらがどこから,どのようにしてやって来るのか,わかっていないのである.

図1:マアナゴConger myriasterのレプトセファルスから稚アナゴにいたる変態過程.発育段階の進行と共に色素発現が進む. (a) 体側に色素が全く見られないレプトセファルス,(b) 尾部にわずかに色素が発現したレプトセファルス,(c)(d) 完全に体側の色素が発現した状態, (e) 変態直後の稚アナゴ(シラスアナゴ),(f) 色素がよく発達した稚アナゴ(クロコ).
2000年晩秋,学術研究船・淡青丸(たんせいまる,610トン)は,マアナゴの産卵場を求めて,東シナ海の陸棚縁辺部に沿ってレプトセファルスの分布調査を行った(Miller et al. 2002). 同じConger属(クロアナゴ属)と見られるレプトセファルスが何個体か採集された.しかし船上の形態的観察では,マアナゴの特徴とされる1列の黒点を体側上にもつ個体は,1個体も見つからなかった(図1の c).
その後研究室に帰って,ミトコンドリアDNAの16S rRNA遺伝子1,217塩基を解析したところ,遺伝子解析用にとりわけていたクロアナゴ属サンプル25個体のうち,20個体はマアナゴ成魚の塩基配列と同じ塩基配列を示した(Ma et al. 2007).これはこの種の特徴とされている体側色素は,これまでの理解とは異なる発達過程を経ることを示している。つまり,全長が74mmより小さいときには色素はみられず(図1の a, b),その後125mm程度に成長して沿岸域にやってくるまでの間に,体側の色素列ができてくるものと考えられる(図1の c, d).
またクロアナゴ属のレプトセファルスの遺伝子解析から,意外なことがわかった.大きいサイズ(60 ? 74 mm TL) のマアナゴのレプトセファルスは東シナ海の調査海域の北部に出現し,小さいサイズ(22, 45 mm TL) は,台湾東方の外洋に出現したのである(図2).これはマアナゴの産卵場はこれまで考えられていた陸棚周辺よりも,ずっと外洋の北赤道海流域にあることを示している.北赤道海流域に分布する小サイズのレプトセファルスは,やがて台湾沖で黒潮に入り,大サイズの分布する東シナ海へと運ばれるらしい.
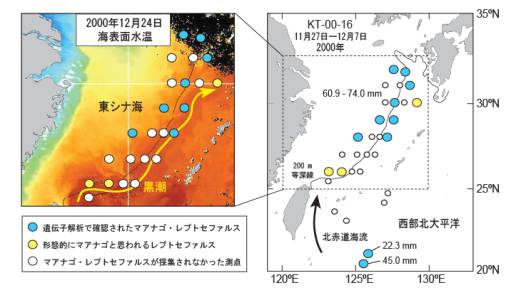
図2:2000年11-12月,東シナ海と西部北太平洋で実施された淡青丸KT-00-16航海で採集されたマアナゴレプトセファルスの分布.北赤道海流(NEC)の水は黒潮に流れ込み,東シナ海を通過する.航海のすぐ後に観察された表面水温は(左図),黒潮の流路(濃い赤色部分)を示しており,観測点はこれに沿って配されている.
遺伝子解析を導入することにより,マアナゴの産卵場が外洋にある可能性が示された.さらに小型のレプトセファルスを外洋で探していけば,いずれマアナゴの真の産卵場に行き着くものと考えられる.その回遊経路は,同じく北赤道海流域で産卵するニホンウナギに比べればやや短いものの(Tsukamoto 1992, 2006; Tsukamoto et al. 2003),北赤道海流と黒潮を利用する点では基本的に同じである.また北大西洋の北アメリカ大陸沿岸に分布する同属のアナゴConger oceanicus(McCleave and Miller 1994) も外洋で産卵することが知られている.今回の発見により,日本のマアナゴもニホンウナギや他の温帯ウナギと同様,長い産卵回遊をすることがわかっている数少ない種のひとつに仲間入りした.
この外洋におけるマアナゴレプトセファルスの発見と体側色素の発達過程を報じた論文は,Fisheries Science 誌に掲載され(Ma et al. 2007),その研究成果に対して日本水産学会から平成19年度Best Paper Award が与えられた。
参考文献
岡村明浩,宇藤朋子,張寰,山田祥朗,堀江則行,三河直美,田中悟,元信堯,岡英夫.2000. 渥美半島太平洋岸におけるマアナゴ成熟度の季節変化. 日本水産学会誌. 66:412-416.
Kimura Y, S. Ishikawa, T. Tokai, M. Nishida, and K. Tsukamoto. 2004. Early life history characteristics and genetic homogeneity ofConger myriaster leptocephali along the east coast of central Japan. Fisheries Research. 70: 61-69.
Kurogi, H., N. Mochioka, Y. Takai, and O. Tabeta. 2002. First offshore record of Conger myriaster leptocephali in the East China Sea. Fisheries Science 68: 1155-1157.
Ma, T., M. J. Miller, J. Aoyama, and K. Tsukamoto. 2007. Genetic identification of Conger myriaster leptocephali in the East China Sea region. Fisheries Science 73: 989?994.
McCleave, J.D., and M.J. Miller. 1994. Spawning of Conger oceanicus and Conger triporiceps (Congridae) in the Sargasso Sea and subsequent distribution of leptocephali. Environmental Biology of Fishes 39: 339-355.
Miller, M. J., T. Otake, G. Minagawa, T. Inagaki, and K. Tsukamoto. 2002. Distribution of leptocephali in the Kuroshio Current and East China Sea. Marine Ecology Progress Series 235: 279-288.
Tsukamoto, K. 1992. Discovery of the spawning area for the Japanese eel. Nature 356:789-791.
Tsukamoto, K. 2006. Spawning of eels near a seamount. Nature 439: 929.
Tsukamoto, K., T. Otake, N. Mochioka, T.W. Lee, H. Fricke, T. Inagaki, J. Aoyama, S. Ishikawa, S. Kimura, M.J. Miller, H. Hasumoto, M. Oya, and Y. Suzuki. 2003. Seamounts, new moon and eel spawning: the search for the spawning site of the Japanese eel. Environmental Biology of Fishes 66: 221-229.
![]()